皆さんこんにちは。
図書館学習サポーター自然研M2のN.Dです。周りの先生方に恵まれ、1つ目の論文を出すことができましたのでここで解説記事を書きたいと思います。論文へのリンクは以下の通りです。
~図書館職員メモ~
この論文が掲載された雑誌Archives of Virologyは、新潟大学が契約するProQuestのコレクションに収録されています(1 year embargo)。
私は普段ユリや野草に感染するオオバコモザイクウイルス (PlAMV) を題材に研究しています。近年、植物ウイルスによる新規病害が数多く発生する中で、野草のウイルスが発生源になっているのではないかと推測されており、野草のウイルスの性質を調べることが重要視されつつあります。
2003年に青森県の野草タチツボスミレから分離されたPlAMV Vi分離株には緑色蛍光タンパク質 (GFP) が組み込まれており、感染した植物組織が緑色に光ります。
たとえ風変わりな植物から分離されたウイルスであっても「ベンサミアナタバコ」や「キノア」という植物ウイルス関連の実験でよく用いられる植物には感染可能な場合が多いため、まずはこれらの植物にウイルスを接種してみました。
すると、このウイルス (PlAMV Vi-GFP) を接種した植物は植物体全身でぼんやりとしたGFP蛍光が観察されたことから、このウイルスは感染性はそこまで強くないながらもベンサミアナタバコやキノアに無病徴で全身感染することが分かりました (図1)。

この結果を受けた当時の私は、「野草由来の病原性の弱いウイルスが、植物に病気を引き起こすポテンシャルを調べれば、将来的に農業的な意味が生まれるかもしれない」と直感で思いました。
そこで、ウイルスの汁液接種と28日間の育成を繰り返して14代に渡る長期継代を行い、実験室内でウイルスを進化させて明らかな病気を引き起こすウイルスの出現を待つ実験をやりました (学部4年~M1まで約1年かかりました)。
PlAMV Vi-GFPをタバコで数ヶ月間継代していたところ、葉がしわしわになってしまう変異ウイルス (P5 mutant) が出現しました (図2)。これを私は「縮葉変異株 (Leaf Crinkle line)」と名付けて日々愛情を注いでいます。

P1-P14はウイルス継代各代の植物、P5 mutantは縮葉病徴が観察された植物、Bright-fieldは室内灯下で撮影した植物、GFP fluorescenceはウイルス由来のGFP蛍光を示す。
別の植物体における縮葉病徴の再現性を確認するために、縮葉変異株を健全なタバコに感染させると、安定して葉がしわしわになる病徴が観察されました。
この結果から、この縮葉病徴 (=しわしわ病徴) はどうやらウイルス感染によるものだということが分かりました。

Bright-fieldは室内灯下で撮影した植物、GFP fluorescenceはウイルス由来のGFP蛍光を示す。
継代後のウイルス変異株はウイルス由来GFP蛍光が強く、タバコへの感染性が強くなっているように見えました (図2、図5上)。
そこで、ウイルス蓄積量を経時的に解析することによって、ウイルス接種から全身感染に至るまでのスピードの違いを論文の図のフォーマットに落とし込めたら面白いだろうと考えました。当時思いついた実験系がこちらです。
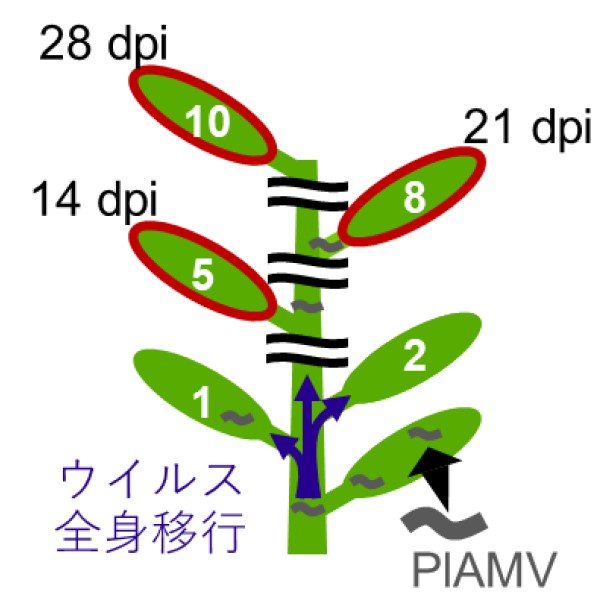
接種後14 日 (14 dpi) における接種葉から5 枚上葉、接種後21 日 (21 dpi) における接種葉から8 枚上葉、接種後28 日 (28 dpi) における接種葉から10 枚上葉のウイルス蓄積量をRT-qPCR を用いて解析することによってウイルスの全身移行・蓄積タイミングを調査した。
この実験を行った結果、継代前と比較して無病徴のまま継代された継代14代目株は約1週間早く、縮葉変異株は約2週間早くウイルス接種葉から上葉に移行・蓄積することが明らかになりました (図5下)。

接種葉からのウイルスの全身移行・蓄積を上葉で待ち構えて、ウイルス蓄積量を経時的に定量した。上側は植物の観察、下側は経時的にウイルス相対蓄積量を解析した結果を示す。異なるアルファベットは有意差を示す。
最後に継代後のウイルスゲノム全長を読んで継代前と比較しました。具体的な変異はブログに載せませんが、縮葉変異株特有の計7箇所の変異が同定されました。実際の論文には継代後のウイルスの変異情報が載っています。
これらの実験結果をまとめると以下のようになります (図6)。

同一の植物で数代感染を繰り返すだけでこんなに性質が変わってしまうなんて、本当にこのウイルスには驚かされてばかりです。
本研究において、単一植物種での継代によって病原性が激化したということは、もしかしたらモノカルチャーな圃場環境が新しい植物ウイルス病の発生を助長しているのかもしれません。
これらのウイルスの性質を決定付ける変異箇所については、卒業までに明らかにすることが出来ませんでした。なんとも悔しい限りですがこの研究を引き継いでくれる後輩を応援するしかありません。
また、地味ではありますが、この研究成果で2023年の農学部主催の国際シンポジウムでポスター賞を頂くことができました。支えてくださった方々に感謝申し上げます。

私は修士課程修了後は研究から離れますが、優秀な研究者・大学院生の方々に私の研究について知ってもらい自身の研究に活かして頂ければ良いのかなと思っています (それ故この記事を書いています)。
最後に、共同研究でお世話になった方々、学会発表や中央図書館のライブラリーラウンジで行われた院生会の研究発表会を聞きに来てくださった方々、農学部生物資源科学プログラムの植物分野の方々、そしてこのブログを読んでくださった方々が良い研究成果を出せるように祈っております。
研究は諦めかけてからが勝負だと僕は思っています。
上手くいかない時でも粘り強く頑張ってみてください。